人类对重要栽培植物的驯化历史总是充满好奇。长期以来, 研究者利用各种技术和方法千方百计地追溯它们的进化历史, 获得了丰硕的成果(Doebley et al, 2006; Hancock, 2012)。这些研究不仅加深了我们对这些物种进化历史的理解, 从而能更好地保护和利用这些重要物种的遗传资源, 而且也使我们更加全面地理解人类文明的发展历史。同时,研究栽培植物驯化起源与进化建立起来的理论和方法也极大地推动了进化生物学的发展(Marcussenet al, 2014)。
茶是最为古老和全球最受欢迎的无酒精类饮料, 它不仅对中国人的文化、健康、医药、贸易等产生了深远的影响, 而且在亚洲乃至整个世界众多人的日常生活中也发挥着重要作用, 茶树已成为许多地区有重要价值的栽培植物。100多年来, 栽培茶树的起源和进化同样吸引了众多研究者的关注(Baildon, 1877; Wright, 1959; 吴觉农等, 1979; 朱自振, 1982; 陈进和裴盛基, 2003; Huang et al, 2014;Meegahakumbura et al, 2016; Yang et al, 2016), 尽管已获得一些重要进展, 但远不能与人类对小麦(Triticum aestivum)、玉米(Zea mays)等作物的栽培历史的认识相比, 依然存在许多争议和谜团。本文回顾和总结了有关栽培茶树驯化起源与进化的成果,特别是近年有关茶遗传多样性研究的相关成果, 讨论了尚存在的问题, 并对未来的研究方向提出了建议。
1 野生茶资源的调查及新物种的发现
任何栽培植物都有其野生祖先, 寻找栽培植物的野生类型或野生近缘种是追寻其进化足迹最好和最常用的方法。茶树是我国最早栽培的木本作物之一, 有悠久的栽培历史和广阔的栽培区域, 因此,长久以来几乎无人会怀疑栽培茶树起源于中国。但自19世纪30年代英国人在印度东北部阿萨姆地区发现疑似野生茶树后, 就有人提出印度阿萨姆是茶树原产地的假说。可见野生资源调查在确认栽培植物驯化起源中的重要性。
中国古代文献对野生茶树已有很多记载(陈宗懋, 1992), 但系统、全面的调查还是在中华人民共和国成立后才进行, 各地的茶科所、林业局、农业及林业院校等部门都参与了野生茶树资源的调查。经过数十年的努力, 获得了中国野生茶树资源极为详细的信息, 相关成果见《中国古茶树》(中国茶人联谊会和中国茶叶学会, 1994)一书及一系列文献(陈兴琰等, 1979; 虞富莲, 1989; 郭远安, 1990; 王平盛和虞富莲, 2002; 田永辉等, 2008; 何青元等,2009; 李苑菱等, 2017)。这些研究发现, 在我国不仅西南地区(包括云南、四川、重庆、贵州)分布有较多的野生茶树, 而且在广西、广东、福建、湖南、湖北、江西等地也有野生茶树的分布。尽管大多数工作并未论证这些野生茶树的分类学地位及其与栽培茶的关系, 多以野生型、过渡型和栽培型进行划分, 但大规模野外考察获得的标本与信息推动了栽培茶近缘种或者说茶组植物的分类学研究, 其中张宏达和闵天禄两位先生为此做出了最为杰出的贡献(张宏达, 1981; 张宏达和任善湘, 1998; 闵天禄,1992, 2000)。
1958年英国学者J. H. Sealy在他的山茶属专著A Revision of the Genus Camellia一书中, 只记录了5种2变种茶组植物, 包括茶[Camellia sinensis (L.) O.Kuntze]、大理茶[C. taliensis (W. W. Smith) Melch.]、依洛瓦底茶(C. irrawadiensis Burua)、细柄茶(C.gracilipes Merrill ex Sealy)和毛肋茶(C. pubicostaMerrill), 其中, 茶又分中国茶变种[C. sinensis var.sinensis (C. B. Clarke) Ridley]和阿萨姆变种[C. sinensis var. assamica (Masters) Kitamura]。《山茶属植物的系统研究》一书出版时, 茶组植物已增至17种(张宏达, 1981), 到1998年《中国植物志》49卷(3)出版时, 茶组植物增至32种。其中, 张宏达及合作者发表新种26个, 其模式产地全部位于中国, 特别是中国西南地区(表1)。尽管有些新种可能是已知种类的同物异名, 并在实际工作中也很难区分, 但张宏达等的工作还是极大地推动了我们对栽培茶野生近缘种的认识, 成为研究栽培茶起源的重要基础。
在上述工作基础上, 闵天禄(1992, 2000)先后两次对茶组植物进行修订, 根据花柱离生、苞片宿存将毛肋短蕊茶(C. pubicosta Merr.)从茶组分出, 并将其他茶组植物归并为12种10变种(表1), 其中老挝茶(C. sealyama Ming)为新种。闵天禄先生的修订一定程度上澄清了茶组植物种类划分过细的问题,Flora of China采用了闵天禄的系统。按照这一分类系 统 , 广 泛 栽 培 的 茶 树 属 于 C. sinensis (L.) O.Kuntze下的两个变种: 阿萨姆变种(俗称大叶茶, 下同)和中国茶变种(俗称小叶茶, 下同); 除此之外,还有多个种或变种也被少量栽培, 包括大理茶、大苞茶(C. grandibracteata Chang et Yu)、德宏茶[C.sinensis var. dehungensis (Chang & B. H. Chen)Ming]、白毛茶(C. sinensis var. pubilimba Chang)、毛叶茶(C. ptilophylla Chang)等。值得注意的是在中南半岛和南亚还栽培一种特殊类型的茶, 即高棉茶或柬埔寨茶(Cambod tea), Wight (1962)将其视为大叶茶的一个亚种[C. assamica ssp. lasiocalyx (Watt)Wight], 其形态特征介于大叶茶和小叶茶之间, 但这一名称在张宏达先生和闵天禄先生的系统中都未进行处理。最近Das和Ghosh (2016)对这一类型进行了分类修订, 将其定为 Camellia sinensis (L.) O.Kuntze var. lasiocalyx (G. Watt) A. P. Das & C.Ghosh。但是, 基于遗传结构的分析证明这一类型的茶实际上是小叶茶和大叶茶之间的杂交品种, 并不是一个自然的分类群(Meegahakumbura et al, 2016;Wambulwa et al, 2016a)。值得注意的是在那些少量栽培的种类中, 大苞茶和大理茶的模式标本就采自村寨附近的栽培个体, 毛叶茶则是近年才开始被有规模地栽培。除上述类群外, 野外调查中还见到民间采集多种野生茶树的叶制成茶, 但规模都不大,包括大厂茶(C. tachangensis F. C. Zhang)、厚轴茶(C.crassicolumna Chang)以及广西茶(C. kwangsiensisChang)等。近年随着市场上鼓吹大树茶的品质更好,从野生或半野生乔木型茶树上采集叶片来加工成茶的现象明显增多, 对茶树资源造成了巨大破坏(Zhao et al, 2014; 李苗苗等, 2015)。因此, 当我们讨论栽培茶的起源时, 首先需厘清到底讨论的是哪一类茶, 同时也需厘清所涉及的野生茶树到底属于哪个物种。
2 栽培茶的起源
祖先类型、起源地点、起源时间以及驯化历程是研究栽培作物起源的4个关键问题, 解决这些问题往往需要多个学科共同努力。有关茶起源的研究很多, 但限于篇幅, 本文仅讨论文献学、语音学、遗传学和考古学4个方面的进展。
2.1 有关栽培茶起源的文献学证据
尽管中国存在最丰富的古代茶文献, 然而汉代以前的相关文献还是很少, 更没有栽培茶在何地何时起源的记载。众多学者已对这些文献做了全面的论述(参见姚国坤和程启坤, 1992), 认为秦汉时期涉及茶的可靠而又明确的记载只有以下4则: 《尔雅》、司马相如的《凡将篇》、王褒的《僮约》以及杨雄的《方言》。至于“神农尝百草”发现茶的药用效果目前还属于传说, 《华阳国志·巴志》提到巴国向周武王进贡的“荼”以及《尚书·顾命》中的“王三宿, 三祭, 三咤”中的“咤”是否就是茶也还有待确认。
古代文献常常充满争议, 但以下几个事实应该是明确无误的: (1) “茶”这一植物或物品在古代汉文字中存在多种多样的写法, 包括荼(cha, tu)、苦荼、槚(cha, jia, gu)、茗、荈、蔎(she)、蕸、荈诧, 但肯定不写作“茶”, “茶”字出现在唐朝, 在陆羽的《茶经》出版后方广为使用; (2)最早的三条有关茶的可靠记载都发生在蜀地; (3)在汉字体系中, “荼”和“槚”这一草一木两种植物最早肯定不是指茶这种植物, 在西汉或更早才开始用来表示茶。正是根据上述文献以及其他明确记载巴蜀之地有野生乔木型茶树的古代文献, 有学者推测巴蜀之地是栽培茶的起源地(朱自振, 1982)。我们认为这些文献记载或许还无法确凿地证明上述结论, 但无疑可证巴蜀之地是最早规模化栽培和饮用(食用)茶的地区之一。现代考古也证实上述文献记载所言非虚, 西汉确已有茶(Lu et al, 2016), 这将在后文论述。汉以后, 有关茶的记载越来越多, 其中详细记录了巴蜀是第一个茶生产中心, 自三国、两晋至唐,茶的栽培中心从西向东移, 至宋、元再向东南移动的过程。对此, 已有学者做过较为全面的综述(见朱自振, 1992), 这里不再赘述。
2.2 栽培茶起源和传播的语音学证据
古代许多族群缺乏文字, 因而记载茶的古老文献很少, 但所有族群都有语言, 而且不同族群通常语言不同, 即使同一族群也会有不同的方言。因此,从不同区域的人们如何说茶这一植物或物品的信息中, 可以追溯茶文化的起源、传播与发展, 众多学者已经做出了有益的尝试(高晓山, 1984; 周靖民,1985; 李锦芳, 2005; 薛德炳, 2015)。这不仅涉及茶本身的使用、栽培和制茶知识的传播, 还涉及民族的迁移、融合等。近代茶在欧洲、非洲等地区的传播及茶发音的演变为追溯古代茶在国内及附近地区的传播提供了重要的参考和启示, 因为近代的传播过程有详细的文献记载可以确证。如在中国以外的地区, 特别是几乎包括印度的所有地区, “茶”存在两类不同的发音: tea类和cha类。前者可从福州、潮州方言追溯至厦门方言(te/tay), 后者可追溯至广州一带的方言(cha) (高晓山, 1984; 马晓俐, 2010),这是中国最早对外开放的两个重要口岸, 显然也是中国最早向西方输出茶叶的地区。
那么厦门的te/tay和广州的cha的音又来自何处呢?各自独立出现还是存在关联涉及茶在中国的起源。“cha”的音分布很广, 哪里的人最先说这一音的呢?有学者推测最早的cha音来自古代巴人(薛德炳, 2015), 认为巴蜀地区至今有多种发音, 包括cha、she、jiameng、chuan cha等, 且与古老方国的“苴” (多音, 发ba, cha等)相关联, 至今川东、渝东的居民饮浓茶时, 仍有ba chu, ba sha, ba che等语音(薛德炳, 2015)。
但也存在另一种可能, 这一发音来自南方的其他族群, 其中之一就是云南南部的孟高棉民族(被称为北方孟高棉族)。现阶段属于孟高棉语系的族群主要分布于中南半岛, 在我国云南分布有佤、德昂(崩龙)和布朗3个民族(马曜, 1978), 后两个民族被认为是最早使用和栽培茶的民族之一。这3个民族都没有文字, 其中佤族将茶说成cha或la, 布朗族把茶说成la, 德昂族则说成jaju (李锦芳, 2005; 李全敏,2013; 王仲黎, 2016)。特别值得注意的是后两个民族的发音和意义, 德昂族中ja意为祖母和外祖母,而ju意为眼睛亮了, 传说是茶治好了古代德昂族王子母亲的眼疾(李全敏, 2013); 而布朗族语言中茶的发音la33 (中平调)与普通叶子的发音la51 (高降调)很近。这表明德昂族的ja以及布朗族的la33 (中平调)的发音很可能并不是来源于茶(cha)的直接转用, 甚至可能其他地区茶“cha”的发音最早就来自这两个民族。布朗族的另一个发音似乎支持这样的猜测,布朗族将茶还用作食材, 是布朗族先民的“野菜”之一, 这种野菜常用作佐料, 布朗族称之为“得责”[tɤ55 tɕiat31], 与其他地区的任何一种有关茶的发音都不相似, 暗示了这些民族或许是独立获得这些知识。德昂族与茶的密切联系还可以从该族的史诗《达古达楞格莱标》得以印证: 在这部长达558行的史诗中, 德昂族以茶为万物之源, 把茶树当作万物的始祖, 人类的祖先, 祖祖辈辈都把茶树视作具有生命、意志和伟大能力的对象而加以崇拜(赵腊林和陈志鹏, 1981; 周灿和赵燕梅, 2013), 这在使用茶的各民族中独一无二。尽管上述史诗描绘了德昂族与茶的密切联系, 但目前还不知道这一史诗产生于何时、何地, 因而也就不能明确判断德昂族何时与茶建立如此密切的联系, 相信随着对该史诗的深入研究, 会得到更为清晰而明确的信息。
我国华南地区一些讲侗台语的民族对茶还有一些十分不同的称谓, 如“过罗”、“拘罗”、“瓜卢”等, 这些发音听起来与“cha”十分不同, 但据李锦芳(2005)研究, 它们实际上与茶的早期发音“槚”、“皋卢”密切相关, 认为后者就来自古代南方百越(侗台)语。这样看来, 国内所有茶的发音都存在高度的相似性和相关性。这一结果暗示了早期茶知识来源的单一性及茶知识传播的方向。
根据上述信息, 我们虽不能确定是华南(古代侗台族群)还是云南南部(古代北方孟高棉族等)或是古代巴蜀中的哪一地最先获得使用和栽培茶的知识, 但可以断定的是无论哪一个族群最先获得茶的知识, 相互之间肯定存在非常早的交流。现有的考古发现表明巴蜀之地不仅与长江中下游地区存在密切的交流, 与云南以及中南半岛北部地区也存在久远和频繁的交流, 现在生活于中国南方以及中南半岛北部的多个民族就是原生活于长江流域的两个古老族群百濮和百越南迁的直接后裔或混血后代。如上文所述的云南孟高棉族群(佤、德昂和布朗), 他们的起源虽还众说纷纭, 但研究表明即使他们不是濮人的直接后裔, 也肯定受到南迁濮人部落的影响(何平, 2003; 李晓斌等, 2007)。从巴蜀至云南南部的广大地区都分布有多种多样的茶组植物,只要拥有栽培和使用茶的知识, 有理由相信这些知识将随族群的迁移而流传, 并在族群迁移中就地取材当地的野生茶树加以利用。大理茶的栽培很有可能就是获得茶知识的当地民族独立开展的。如果没有茶的知识, 很难想象大理茶分布地的民族也会将这一物种发与茶相似的音(如白族将茶发成tso)。
景颇族对茶的发音也是值得研究的, 英国人Robert Bruce于1823年在印度阿萨姆地区得到的茶树就是当地景颇族所种植 (Taknet, 2002)。景颇族不是当地的土著民族, 该民族的记忆及考证表明他们来自北方青藏高原的东部地区, 几经迁移到达现在的云南、缅甸和印度东北部。可能早在汉代这一民族就进入云南境内, 至宋代, 西进的景颇语支系可能沿独龙江南下到恩梅开江流域与缅语支等汇合,成为现代景颇族分布的雏形(李怀宇, 2003)。如果说这一民族没有在南迁过程中学会利用、栽培茶的知识是很难令人相信的, 因为他们迁移所经过的地区早已有大量茶栽培。但景颇族称茶为pha lap (李锦芳, 2005), 这听起来与cha、tsa等发音差别很大, 为何如此, 还有待研究。
2.3 有关栽培茶起源的遗传学证据
遗传物质是追溯一个分类群谱系发生最直接的证据。随着分子标记技术的发展, 自20世纪末,中国、印度、日本等国的学者利用多种分子标记技术分析了不同区域、不同栽培茶的遗传多样性和遗传结构, 本文列举了其中最主要的28项研究(表2)。这些研究收集的样品来源不同、数量不一、使用的分子标记也各不相同, 因而结果很难一一比较。尽管如此, 分析这些研究, 我们至少可以得到如下重要信息, 有助于更深入地认识茶树的驯化起源和传播。
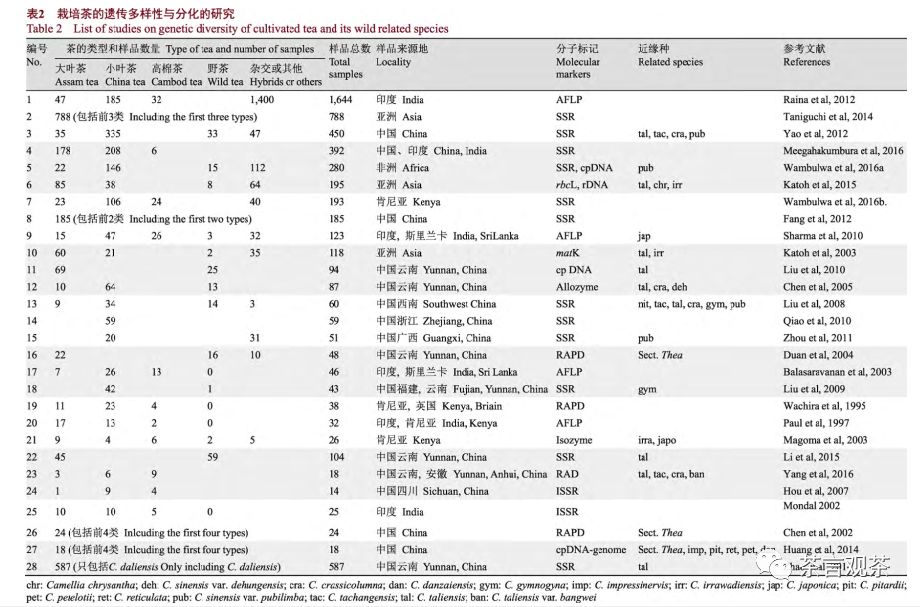
(1)在人类作用下, 茶品种间的杂交、基因渐渗以及栽培茶与野生近缘种杂交和渐渗十分普遍。例如, 高棉茶很可能就来自大叶茶与小叶茶的杂交(Meegahakumbura et al, 2016); 而印度栽培茶的品种十分混杂, Raina等(2012)对印度1,644份栽培茶样品的遗传多样性的研究也表明, 其杂交类型十分丰富, 并且发现遗传类型与基于形态特征的分类型并不一致, 印度很多栽培茶为印度产大叶茶和中国产小 叶 茶 之 间 的 杂 交 品 种 (Meegahakumbura et al,2016); 非洲栽培茶的历史不到200年, 但其栽培茶群体中存在多个类型且品种间遗传差异大, 主要包括来自中国的小叶茶和印度的大叶茶以及它们间的杂交品种, 且多以大叶茶为母本的F1代和回交后代的杂交品种为主(Wambulwa et al, 2016a, b, 2017);对大理茶(包括C. irrawadiensis Burua)和中国大叶茶居群的遗传结构分析则发现大理茶的遗传物质渗入到大叶茶中, 暗示了大理茶很可能参与了大叶茶的驯化过程(刘阳等, 2010; 李苗苗等,2015)。
(2) Meegahakumbura等(2016)利用SSR标记, 通过分析来自中国和印度的392份茶树种质资源(不仅包括了大量古茶树和近似野生茶树的样品, 还包括了两个国家主要产茶区的茶树品种资源)的遗传组成与结构, 结果表明, 栽培茶树可分为3个遗传谱系, 代表了3种不同的茶树类型, 即小叶茶、中国产大叶茶(主要产云南南部和西部)和印度产大叶茶,后两者虽同属大叶茶, 但代表不同的遗传谱系, 进而推测小叶茶、中国大叶茶和印度大叶茶很可能各自独立起源, 中国大叶茶很可能起源于云南西南(南部及西部地区)及相邻区域(如缅甸), 而印度大叶茶则可能起源于印度阿萨姆地区。对叶绿体DNA片段的分析也表明印度阿萨姆地区确实栽培着一类谱系上与中国大叶茶十分不同的茶树(Meegahakumbura, 2016)。
(3)悠久的茶树栽培区存在众多地域性类型,这些地方品种的遗传组成与其他区域的品种十分不同。除了上文所述的中南半岛的高棉茶和印度的大叶茶外, 日本的栽培茶也十分不同, Taniguchi等(2014)利用SSR标记分析了日本从亚洲各国范围内收集的788份茶种质资源的遗传组成, 结果发现可以将这些茶分为3类。 其中, 日本栽培的品系构成独立的一类, 但遗传多样性低, 来自其他区域的茶构成另外两类, 分别对应大叶茶和小叶茶; 中南半岛各国及孟加拉、斯里兰卡以及印度的部分品种似乎属于前者, 而中国、韩国、印度的很多品种属于小叶茶。日本的栽培茶叶无疑最早是从中国引入的(Matsumoto et al, 2002), 但上述结果表明长期的人工选择使日本的栽培茶与中国小叶茶发生了遗传分化, 两者间的遗传差异甚至比大叶茶与小叶茶的遗传差异还要大。而根据Katoh等(2003)的分析, 缅甸栽培的茶树遗传类型也十分丰富, 不仅包括大叶茶、小叶茶, 还有些来源于大理茶(包括C.irrawadiensis Burua)等。
(4)中国的栽培茶拥有十分丰富的遗传多样性和栽培类型, 除了上文所述的小叶茶、大叶茶外,在云南西南地区, 长期以来当地居民也栽培大理茶、大苞茶和德宏茶(Zhao et al, 2014)。Yao等(2012)分析了采自中国各茶区的450份茶树样品的遗传结构, 这是到目前为止对中国栽培茶遗传多样性最全面的研究, 不仅包括的样品多和广(包括331份采自中国各茶区的地方品种, 87份引入品种以及大理茶、大厂茶、厚轴茶、白毛茶等32个野生、半野生类型), 而且所检测的EST-SSR位点多达96个。结果发现: ①地域性的分化十分明显, 从遗传结构上看可以分成明确的5类, 湖南以东(包括湖南)的长江下游各省以及福建的栽培茶遗传上高度一致, 分为一类, 广东与广西、贵州与重庆、湖北与四川相邻两省的栽培茶品种除少数个体外, 各有十分相似的遗传组成而分别分成3类, 云南的地方品种独成一类;②云南、贵州、广西三省的茶有较高的遗传多样性; ③按遗传组成区分的5类茶, 并不与分类学上的种或亚种对应, 小叶茶和大叶茶与野生种类的关系并未得出结论, 尽管湖北、四川、重庆、贵州4省栽种的地方品种都属于小叶茶, 但其遗传组成上与长江下游的小叶茶并不一致(Yao et al, 2012)。
(5)利用RAD-seq技术, Yang等(2016)分析了包括大叶茶、小叶茶、大理茶、厚轴茶等18个茶组植物个体的遗传关系, 研究材料中包括从形态特征上被认为是介于野生与栽培的中间过渡类型的邦威大树茶。结果表明邦威个体确实不同于大叶茶与小叶茶, 也不同于大理茶等其他种类, 是前两类栽培茶最近的姐妹群, 这一结果支持栽培的大叶茶和小叶茶都来自云南南部的假说, 但还需要更多的材料和数据进行证实(Yang et al, 2016)。实际上, 表2所示的研究结果彼此之间的不一致更为突出, 限于篇幅不能一一讨论。这里只举一个突出的例子。Huang等(2014)测定了13种(含亚种)的叶绿体基因组序列, 根据其结果得出如下结论:
(1)大叶茶和小叶茶不仅是独立的起源, 而且是完全不同的种, 前者的祖先与毛肋茶是姐妹种, 后者则与膜叶茶(C. leptophylla S. Y. Liang et Chang)关系密切, 而白毛茶也为独立的种; (2)从叶绿体谱系上看大理茶甚至不属于茶组。这一结果无疑是令人吃惊的, 从形态特征上看, 大理茶无疑是茶组植物, 毛肋茶、膜叶茶与小叶茶也区别明显, 特别是毛肋茶因有离生的花柱和宿存的小苞片甚至被归入离蕊茶组(闵天禄, 2000)。上述结果与根据等位酶和部分叶绿体基因进行的群体遗传分析获得的结果也相矛盾, 例如有研究表明大理茶与大叶茶十分相似,谱系上甚至不能分开(Chen et al, 2005; 刘阳等,2010; 李苗苗等, 2015) (表2)。根据DNA信息研究茶组植物的遗传多样性及分类系统, 结果为什么会有如此大的差异?一方面,我们认为利用不同的分子标记可能会带来不同的结果: 早期使用RAPD与ISSR进行研究, 其结果的重复性可能会存在问题; 利用叶绿体DNA (cpDNA,母性遗传)与利用SSR位点(绝大多数位于核基因组内)得到的结果也会出现差异; 而同样是利用SSR位点, 如果不是利用严格遵从Hardy-Weinberg平衡的SSR位点来分析, 不同的SSR位点也会得到不同的结果。另一方面, 我们认为更重要的原因在于不同研究使用的样品的准确性、采集范围及数量存在较大差异(表2)。实际工作中发现, 即使按照闵天禄分类系统(闵天禄, 2000), 整个茶组植物种间差异都很小, 不容易识别, 而且无论是种间还是同种不同品种间杂交亲和性都很高(Raina et al, 2012), 正如表1所述, 人类栽培的茶树包括多个种, 广泛的引种给原来分布区不重叠的种或变种提供了众多的杂交机会, 加上人工杂交育种等活动, 使得目前栽培茶的杂交类型十分常见, 这给样品的确定带来困难,那些形态上看起来十分相似的品种遗传组成上很可能明显不同(Raina et al, 2012), 可想而知在取样的范围和数量有限的情况下, 如果样品的准确性再存在问题, 结果必然会发生偏离。
2.4 有关栽培茶起源的考古学证据
对于栽培作物的起源, 考古发掘的遗物是最直接的证据。有关栽培茶的化石和遗存发现很少, 到目前为止, 明确或较为明确的考古证据仅有2例。最为明确的考古发现来自西汉西安古墓和西藏阿里古墓的随葬品。Lu等(2016)在上述两个埋藏中发现了显然是加工过的茶, 特别是西安古墓中的茶为幼芽所制, 与现代茶十分类似。这一发现不仅证明早在2,100多年前, 在西汉的中心地带已有饮茶的习惯, 而且远在西藏阿里的人们也在使用茶叶, 证实司马相如等对蜀地有关茶的记载应该是可靠的。这一发现使我们有理由推断南方栽培茶及饮茶习惯一定比2,100年前早得多。这些发现还引发了十分有趣的问题: 茶是如何传到偏远而海拔很高(平均4,000 m以上)的阿里呢?是从张骞刚开通的丝绸之路传播还是通过传说中更早的身毒(印度)之路流传呢?如果是后者, 茶在印度的流传就要比现在人们的认识早得多。
另一项有关栽培茶起源的发现出自新石器时代的浙江余姚田螺山遗址, 考古学家们在这一古村落距今约6,000年前的堆积层中发现多个保存完好、疑似山茶的根, 通过木材解剖断定这些根是山茶属植物的根(浙江省文物考古研究所, 2011; 铃木三郎等, 2011), 并通过分析比较遗存根及分布于浙江的多个山茶属植物根的茶氨酸含量, 推测这些遗存的根 就 属 于 茶 这 一 物 种 ( 虞 富 莲 , 2010; 程 启 坤 ,2016)。 根据这些根靠近干栏式建筑的一些带垫板柱坑并生活于人工挖掘的熟土浅坑内, 考古学家们断定这些根应为人工栽培而非野生。尽管上述植物根是人工栽培还是野生遗留依然存在争议(竺济法,2015), 而且也不知道当时人们是如何利用它们的,但这一发现还是暗示, 即使今天没有野生茶组植物分布, 这一区域也可能是栽培茶的起源地。如果这一结果被确认是真的, 那将引出一系列有趣的问题。从遗传多样性的研究来看, 现代长江下游各省,包括福建等地栽培的茶树品种无疑都是西部或西南部传播过来的结果(Yao et al, 2012), 田螺山悠久的栽培驯化活动为什么没有像栽培水稻一样在这一区域流传下来?历史上存在百越诸族多次南迁的事件, 是否这一区域的古人南迁时将有关茶的知识传播到南方?这些问题还有待于更多的研究。
3 未来的工作展望
正如前文所述, 任何一种栽培植物的起源都涉及祖先类型、起源地点、起源时间以及驯化历程等问题。就栽培茶的起源来说, 尽管已经取得了可喜的成果, 提出了多个有意义的假说, 这些假说虽彼此间存在矛盾却各有证据支持。但严格说来, 上述4个问题没有一个得到明白无误的证明, 还需要更多的证据。笔者看来, 未来的工作应该特别加强如下方面的研究:
(1)茶组植物的分类和谱系重建。所有栽培植物追根溯源都来自野生种, 尽管我们探求的问题是栽培茶树甚至只是C. sinensis (L.) O. Kuntze中栽培类型的起源, 但也依赖于整个茶组植物清晰的分类界定及明确的谱系关系。正如上文所述(表1), 栽培茶的野生近缘种分布广泛且种类众多, 而且物种的界定仍存在很大争议。例如, 闵天禄(2010)的系统将所有大规模栽培的茶树类型如C. arborescens Chang &F. L. Yu、C. assamica (Mast.) Chang以及C. dehungensis Chang & B. H. Chen等都归入C. sinensis (L.)O. Kuntze这一名称之下, 而张宏达和任善湘(1998)则将栽培的茶划分为至少6个种: 包括: 茶[C. sinensis (L.) Kuntze]、普洱茶[C. assamica (Mast.)Chang]、狭叶茶(C. angustifolia Chang)、多萼茶(C.multisepala Chang)、大树茶(C. arborescens Chang etYu)和细萼茶(C. parvisepala Chang)。前两个种又分别各包括3个变种。上述两个系统的分类都只根据部分形态特征进行, 而很多形态特征的变异常常不是间断的, 这给分类群的识别带来困难。在我们厘清物种的界限问题之前, 只选其中一个种来研究很难获得正确的结论。
另外一个问题更为突出, 山茶属植物种间杂交比较普遍, 同组的种类更容易杂交, 由于栽培茶树与其野生近缘种(茶组物种)通常同域分布(特别在我国西南地区), 而在我国茶的栽培历史又是如此悠久, 从野生种类向栽培类型的基因渗入是可以预见的(如大理茶的基因就可能渗入到大叶茶之中,刘阳等, 2010; 李苗苗等, 2015), 而人为有意的杂交就更为常见(Raina et al, 2012), 这导致茶的遗传来源复杂, 形态变异多样, 因此基于形态的分类非常困难。另一方面, 从栽培向野生的回归也很常见,一些被界定为野生型的古茶树可能是从茶园中逃逸到野外, 而并非真正的野生茶树, 这给研究茶树栽培类型的起源带来困难。因此, 要揭示这一问题需要将更多的种类考虑进来一起研究。目前大多数研究还只是包括栽培品种(表2), 没有或很少有野生近缘种的材料, 因而还不能进行真正的谱系分析。已有一些工作注意到这一问题并做了十分有益的尝试(陈亮等, 2002; Yao et al, 2012; Huang et al,2014; Yang et al, 2016), 但涉及野生近缘种的类型和数量太少或获得的分子信息有限, 结论的可靠性还需确认。鉴于茶组植物种边界的模糊性以及栽培茶起源问题的复杂性, 未来的研究应该将茶组植物作为一个整体严格按谱系地理学要求取样, 构建茶组植物的谱系发生关系, 在此基础上再讨论栽培茶的起源。
(2)构建谱系的分子标记的选择。Avise (2000)指出真实的谱系发生应该不仅需要来自细胞质基因组的信息, 也需要来自核基因组的信息, 对于像茶组植物这样容易杂交的植物来说更是如此。以往的研究发现山茶属植物的叶绿体基因组片段与其他类群相比变异位点很少, 常用的片段(如matK,rbcL等)难以用于研究山茶属物种的谱系发生, 但近来随着众多山茶属植物(包括多个茶组植物)的叶绿体基因组被测定, 发现了一些变异较快的DNA片段。如Wambulwa等(2016a)利用3个进化速率较快的叶绿体DNA片段对非洲8个国家的栽培茶开展了研究, 结果促进了对非洲栽培茶来源的认识。同样,山茶属植物的基因组数据也有报道(Xia et al, 2017),而转录组数据就更多, 这提供了机会筛选单拷贝或低拷贝的核基因来进行双亲谱系关系的研究。SSR位点被广泛用来研究茶的遗传多样性和遗传结构(表2), 但若使用非中性的SSR位点, 则可能会使结果偏离真实的历史(Taniguchi et al, 2014), 而且检测的位点也常常有限, 不能反映整个基因组的分化。此外, 众多SSR位点变化十分快, 用来揭示现阶段群体的遗传结果十分有效, 但要根据其结果来推测其历史就十分困难和不可靠。基于第二代测序技术开发的新分子标记技术, 如RAD、重测序等, 已经被广泛用于其他栽培作物的研究(Huang et al, 2012;Marcussen et al, 2014)。Yang等(2016)使用RAD技术对18份栽培茶树和野生近缘种进行了研究, 对小叶茶与大茶树的亲缘关系及与茶叶品质相关的基因进行了分析, 获得重要的结果。然而, 由于使用的材料十分有限, 这一方法的应用及其所得出的结论还有待进一步验证, 而且目前这一方法几乎还没有用于栽培茶的群体遗传研究。相信随着这些技术的应用, 将会极大地推动我们对栽培茶进化历史的认识, 特别是随着对更多茶及近缘种全基因组序列的测定, 栽培茶驯化种发生的谜团将会逐步被揭开。
(3)语音学及民族植物学研究。语言是人类文明在时空中传播最为重要的工具。尽管不同地区、不同民族对茶这一物品发音的相似性和相关性暗示了茶文化起源的单一性, 然而, 很多研究还只是基于推测, 未来需要扩大至与茶相关的更多词汇, 使用更为科学的方式来追溯相关语音的变化, 特别是应该结合民族植物学的研究。目前有关茶的民族植物学的研究还非常有限(陈进, 2003; 李昶罕和秦莹,2015), 且缺乏更大范围内和更多民族的比较。语音学和民族植物学的整合比较研究不仅能帮助我们揭开茶文化的起源、传播及演变, 也将有助于我们认识茶的栽培类型的起源。例如, 在我国云南及附近地区的一些少数民族还保留多种多样的利用茶树的方式, 而在越南南部的一些山区至今还保留最原始的喝茶方式, 即从茶树上直接采取新鲜枝叶煮水即喝, 这些信息对于研究茶文化的传播都具有重要的意义。近年有关栽培的香蕉起源和传播的研究方法值得借鉴(Perrier et al, 2011), 特别是有关语音学的部分。该文作者收集了1,100个与香蕉相关的词汇, 利用标准的语言分析比较方法(Joseph & Janda,2003), 构 建词 汇 的 邻接树 (neighbor-joining tree),获得了与香蕉相关的词汇语音变化的信息。与茶相关的词汇十分丰富, 而且地域性很强, 值得用相似的方法深入研究。
此外, 越来越多的证据表明, 早在3,000–4,000年前的三星堆文化时期, 巴蜀地区的文明不仅与长江中下游的文明密切相关, 而且与中南半岛甚至印度都存在远比以往认识要密切得多的联系, 如越南北部发现与三星堆高度相似的牙璋, 而三星堆则发现了众多海贝等南海甚至印度洋的物品(段渝, 1993,2015; 张弘, 2009; 彭长林, 2015)。对于后一条路线,云南就是最重要的通道, 因此, 在考虑茶文化的传播时, 必须基于民族的迁移以及不同地区漫长的交往历史这一重要的背景。
(4)缅甸、越南、印度东北部的野生茶组植物。中南半岛的许多区域(包括越南、老挝、缅甸等的北部地区)以至印度东北部的阿萨姆地区也有野生茶组植物分布, 但缺乏详细的调查。已有研究中来自上述地区的野生样品很少, 而且分类学归属也有待确认。根据目前的研究, 我们可以看出这些地区的样品对于阐明栽培茶的起源十分关键。例如, 有研究推测小叶茶、印度大叶茶、中国大叶茶3类栽培茶很可能独立起源(Meegahakumbura et al, 2016),这无疑需要更多的野生茶树样品来进一步确认; 而根据叶绿体全基因组的研究, 小叶茶和德宏茶的祖先与膜叶茶最近, 大叶茶的祖先则与毛肋茶构成姐妹群(Huang et al, 2014), 前一个野生种目前只在广西、越南交界的很小区域有采集记录, 在越南的分布一无所知, 而后一个野生种只在越南的巴维山有记录, 国内所获样品的可靠性有待确认, 需要更多的样品才有说服力。因此, 无论从发现新的野生茶组植物资源的角度, 还是从研究栽培茶起源的角度都需要对这些地区开展更为深入的调查。
(5)近万年以来的全球气候变化与茶分布区的波动。在化石缺乏的情况下, 常常根据野生近缘种及栽培类型的谱系地理关系来推断栽培作物的起源地。但这样推理的前提条件是在栽培作物起源后野生近缘种的分布格局没有重大变化, 否则很可能得出错误的结论。然而, 上述前提条件常常是不成立的, 越来越多的研究表明近1万年来, 由于全球气候的波动及人类活动的影响, 许多动植物的分布区都发生过重大变化(竺可桢, 1972; 黄璜等, 1998;文焕然, 2006)。最典型的例子就是野生稻(Oryzarufipogon)的分布, 大量化石证明在6,000–7,000年前, 野生稻分布至黄河流域, 长江下游地区很可能是栽培水稻的起源地, 但现在野生稻在这一地区已经消失殆尽(黄璜等, 1998), 根据现存群体来推断栽培稻的起源地必然会引发争议(Fuller et al, 2009;Huang et al, 2012)。茶组植物的分布也发生了变化,唐代的陆羽在《茶经》中曾记载“茶者, 南方之嘉木,一尺、二尺, 遁至数十尺, 其巴山峡川, 有两人合抱者, 伐而掇之”, 现阶段在此区域两人合抱的野生大茶树已十分罕见。而长江下游地区尤其是在浙江余姚, 已经没有野生茶树了, 在田螺山遗址发现山茶植物的根, 说明6,000多年以前, 这里完全有可能分布有野生茶树。实际上, 近在20世纪40年代, 在山东省还发现一株粗有三抱、高约5丈的大茶树(陈兴琰等, 1979), 尽管不知道这一孤立的乔木型茶树的来源, 但至少说明在近代大茶树还能在那里生存。鉴于上述原因, 未来在研究栽培茶的起源时,必须考虑近万年来野生茶分布区的可能变化, 还需考虑在漫长栽培历史过程中各地区栽培品种的更替对区域遗传多样性造成的影响。
综上所述, 中国长江流域及以南地区分布有众多茶组植物, 包括各类栽培茶的野生近缘种; 南方各民族语言中“茶”发音的相似性及相关性暗示了茶知识起源的单一性, 最可能的地区在古代的巴蜀之地或云南南部, 而遗传结构上的特征揭示出栽培茶的起源涉及茶组植物的多个物种, 即使就C. sinensis (L.) O. Kuntze (包括多个亚种)这一物种的栽培类型而言, 也可能有多个起源; 历史文献显示了茶文化在长江流域从西向东的传播和发展, 遗传信息也揭示出从古巴蜀之地至长江中下游遗传多样性的减少, 但考古发现却指出很可能最早的栽培茶就出现在长江下游; 在茶的栽培品种、特别是在茶知识的传播过程中, 当地野生茶组植物的基因渗入至栽培类型中, 或直接用当地野生资源培育不同的栽培类型以及人为的杂交等导致遗传上的复杂性。未来的研究需要从整个茶组植物出发, 利用文化、遗传、地质环境变化、考古等多种方法提供明确的证据。
责任编辑:千鹤茶苗