紫色芽叶茶树是一种叶色特异的茶树资源,目前最有名的、研究最广泛的紫芽茶树品种就属‘紫娟’了。紫色叶片组织富含花青素,而花青素具有强的抗氧化活性的健康功效,因此紫芽茶树资源也受到了格外关注。花青素是通过类黄酮途径在细胞质基质中合成的,最终以糖基化的形式储存在液泡中。而茶树的儿茶素类也是通过上述途径合成的,但是目前对于茶树中儿茶素类和花青素的相关性还不十分清楚:首先一般正常的茶树中大量积累的是儿茶素类而不是花青素;其次,在紫芽茶树品种中,花青素的积累与儿茶素的积累为负相关,这就暗示着它们之间的竞争关系;还有,花青素主要积累在液泡中,而儿茶素类则主要集中于叶绿体和脉管壁;最后,PAP1-like MYB基因在控制茶树中花青素的合成中发挥关键作用,但是在烟草中异源表达该基因,则激活了大部分的类黄酮相关结构基因,导致黄酮醇类和花青素的同时积累,这与其在茶树中的功能不一致。当然这些结果也预示着茶树中特异调控花青素的基因的存在,因此很有必要继续对紫芽茶树中花青素富集的分子机理继续探索。
在本研究中,作者首先以正常品种‘龙井43’和紫芽品种‘紫娟’为材料,对二者叶片组织中的儿茶素成分以及花青素含量进行测定,结果发现‘紫娟’中总儿茶素含量显著低于‘龙井43’的,而总花青素含量却是‘龙井43’的16倍。
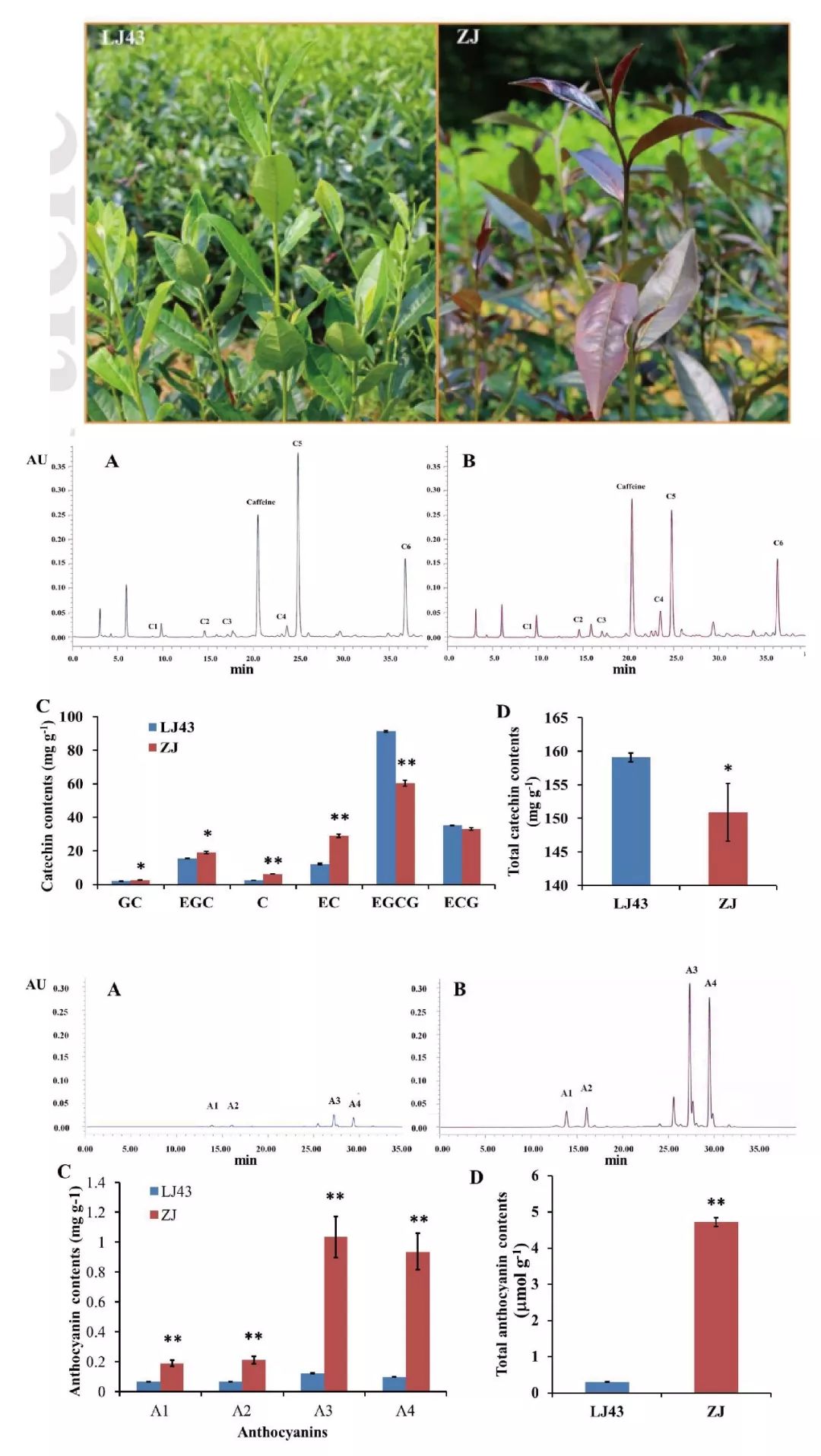
然后作者利用RNA-seq分析二者的差异表达基因,根据MBW转录因子复合体调控花青素积累的事实从差异基因中鉴定到一个PAP1-like MYB基因(CsMYB75)、一个TT8类bHLH基因和两个WD40基因,但是这也不足以解释儿茶素积累差异的表型,虽然花青素合成与儿茶素合成在同一个通路中,但是二者呈负相关关系,这意味着在调控二者积累中必定存在一个下游的通量开关。
接着作者考虑到既然二者在细胞中的分布存在着差异,那么二者的运输系统可能是不同的,因此就集中于与二者积累相关的运输途径最后一步调控基因的表达情况,发现一个F亚组的谷胱甘肽转移酶基因(CsGSTF1)在‘紫娟’中与在‘龙井43’中相比有5倍上调。
对CsGSTF1在‘紫娟’不同组织的表达定量分析发现其表达与花青素的含量正相关,而考虑到GST调控花青素积累主要是调控花青素在液泡中的沉积,因此作者又对两个品种的叶片组织进行显微观察,发现色素主要在叶片上表皮分布,且‘紫娟’的叶片上表皮分布大量的囊泡,而‘龙井43’则很少,预示着CsGSTF1参与花青素沉积可能与活跃的囊泡转运有关。
另外通过花青素含量不同的一系列种质中CsGSTF1的表达分析发现其表达与花青素含量正相关而与儿茶素含量负相关。
然后作者以‘龙井43’ב白毫早’的170个F1单株为材料(这些单株芽和幼叶在夏季呈现由绿到紫的分离表型),基于SSR标记对这个群体构建连锁图谱,将控制叶色变异的QTL定位到LG8,而利用CsMYB75和CsGSTF1基因的特异标记同样将这两个基因定位到LG8。
进一步分析发现这两个基因在叶色表型变异调控效应上不具有显著交互作用,表明二者的等位基因类型与紫芽表型相关性上具有一致性,因此猜测二者可能具有功能相关性,接着通过异源表达实验证实CsMYB75能够强烈激活CsGSTF1的表达;进一步对这两个基因在两个品种中的启动子序列和编码区序列进行比对分析,其中CsGSTF1的基因序列在两个品种中完全一致,而CsMYB75的则存在着差异,因此CsMYB75的序列变异可能在调控叶色变异上发挥着功能。
因为CsGSTF1与拟南芥TT19基因同源性较高,将CsGSTF1在tt19突变体中异源表达却并没有恢复tt19种皮丧失的表型,对转基因株系进行RNA-seq分析发现外源基因CsGSTF1的引入并没有改变拟南芥中花青素合成途径基因的表达,但是却影响了防御响应以及光合作用相关基因的表达。
Abstract
Cultivars of purple tea (Camellia sinensis) that accumulate anthocyanins in place of catechins, are currently attracting global interest in their use as functional health beverages. RNA‐seq of normal (LJ43) and purple Zijuan (ZJ) cultivars identified the transcription factor CsMYB75 and phi (F) class glutathione transferase CsGSTF1 as being associated with anthocyanin hyper‐accumulation. Both genes mapped as a QTL to the purple bud leaf color (BLC) trait in F1 populations, with CsMYB75 promoting the expression of CsGSTF1 in transgenic tobacco. While CsMYB75 elevates the biosynthesis of both catechins and anthocyanins, only the latter accumulate in purple tea, indicating selective downstream regulation. As GSTs in other plants are known to act as transporters (ligandins) of flavonoids, directing them for vacuolar deposition, the role of CsGSTF1 in selective anthocyanin accumulation was investigated. In tea, anthocyanins accumulate in multiple vesicles, with the expression of CsGSTF1 correlated to BLC, but not catechin content in diverse germplasm. Complementation of the Arabidopsis tt19‐8mutant, that is unable to express the orthologous ligandin AtGSTF12, restored anthocyanin accumulation, but did not rescue the transparent testa phenotype, confirming CsGSTF1 did not function in catechin accumulation. Consistent with a ligandin function, transient expression of CsGSTF1 in Nicotiana occurred in the nucleus, cytoplasm and membrane. Further, RNA‐Seq of the complemented mutants exposed to 2% sucrose as a stress treatment, showed unexpected roles for anthocyanin accumulation in affecting the expression of genes involved in redox responses, phosphate homeostasis and the biogenesis of photosynthetic components, as compared with non‐complemented plants.
责任编辑:千鹤茶苗